Greg Albery (Georgetown) presents “Social networks, spatial networks, parasites, and deer"10/29/2019 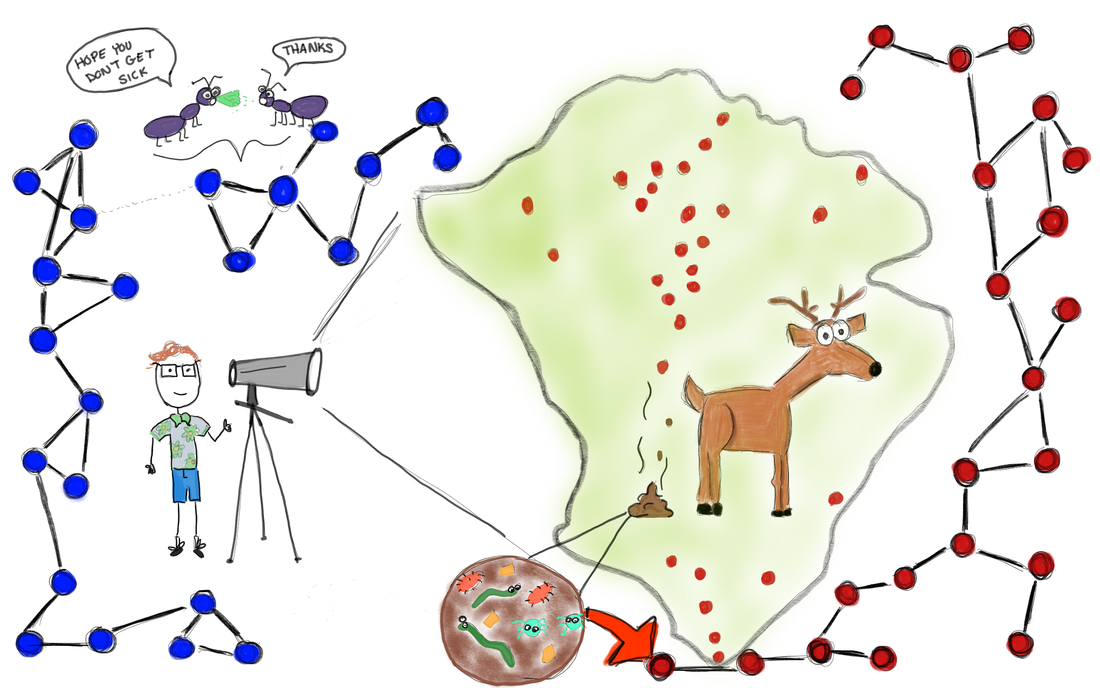 Illustration by Carly Rozins A few weeks ago in the Berkeley EEID seminar we had Dr. Greg Albery present some of his ongoing research. Greg is a recent addition to the Bansal Lab at Georgetown University, but this month he is visiting the Boots lab here at UC Berkeley. Greg’s seminar was on ‘Spocial Networks’. What is a spocial network you might ask? Well this is an important question and a topic often overlooked. But first we need to discuss social networks. A network is a collection of nodes (dots) and edges (lines connecting nodes). Nodes represent hosts and the edges represent interactions, ones that might lead to infectious disease transmission. Networks capture host heterogeneity and provide a rich dataset for determining the effects of sociality on disease dynamics and life history characteristics. Importantly, networks can be empirically derived from surveys, observations, pit tags and radio proximity collars that capture/log social contacts between individuals in the population. These empirical networks are classified as social networks. Therein lies the problem. Since space affects everything, how can these networks be classified as solely social? If space is ignored, it is impossible to determine whether the observed social effects on disease dynamics are actually due to social events, or whether they are truly due to environmental (spatial) constraints. The environment will shape how populations structure and population structure affects social behavior. Greg unraveled these two confounding factors. He collected both spatial and social data for a population of red deer on the Isle of Rum. With an Integrated Nested Laplace Approximation Linear Mixed Models (wha… that was a mouthful!) he determined that spatial heterogeneity is an important factor in both immunity as well as parasitism. Interestingly, he found fine scale spatial patterns in parasitism. Essentially, the best models are spocial, those that control for both spatial and social factors. However, Greg is not done yet! He is now working with an incredibly detailed ant data set on social and spatial behavior and he is on the lookout for even more data. So, if you know of any datasets that include any combination of social, spatial and disease data, Greg would like to chat with you! Summary by Carly Rozins
15 Comments
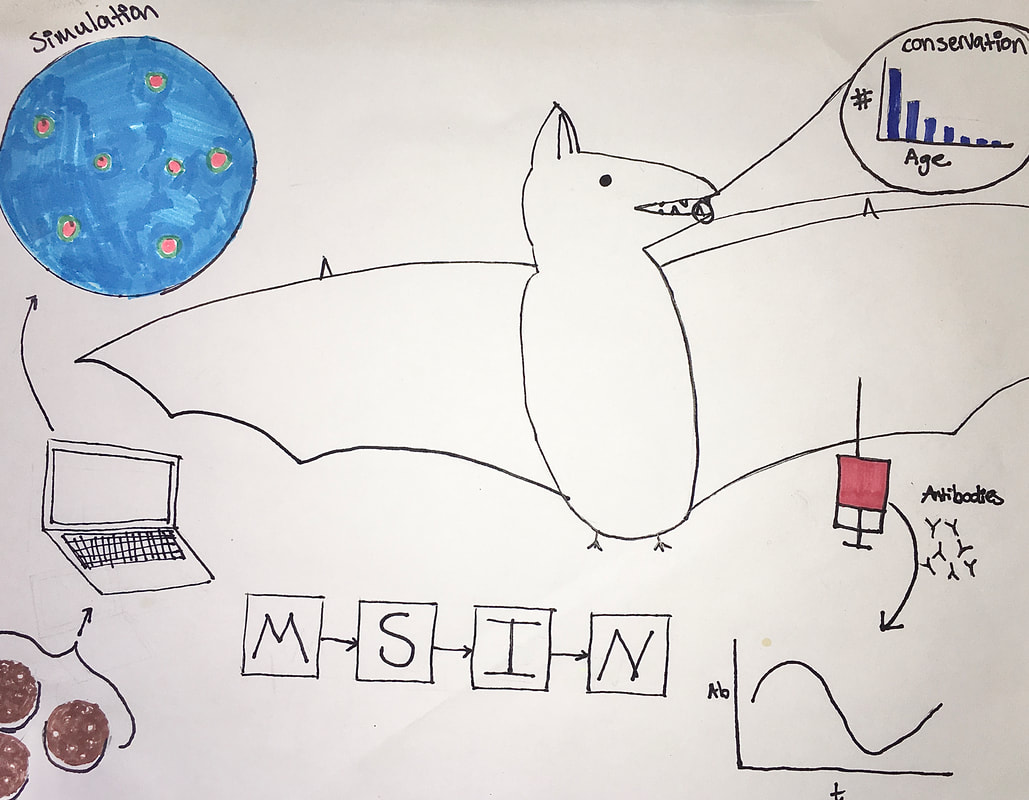 Illustration by Laura Alexander Bats play host to some of the gnarliest, deadliest viruses known today, including those that cause Ebola, Marburg, SARS and MERS. So what gives? Are these fuzzy flying mammals special in some way in their propensity to produce zoonotic pathogens that infect us humans? There’s a growing consensus that the answer to this question is almost certainly “yes!”, but what is it that makes bats special viral vessels? This week, Cara Brook, a Miller Postdoctoral Fellow at UC Berkeley, shared her research seeking to answer this question. Cara started with some basic bat biology that helps explain why they may be so special. Bats are among the longest-lived mammals for their size, are highly gregarious, do that whole flying thing, and have some curious immune functions. Among these are partial resistance mechanisms to viruses that help prevent pathology such as inflammation that is normally associated with viral infection. Even more unique is their constitutive expression of IFN-, an anti-viral cytokine. This suggests that bats are better equipped than other mammals to fight off viral infections, and do so with minimal associated pathology! Seasonality in seroprevalence and viral shedding rates among bats suggests that there may be more to these anti-viral immune responses, though. Cara’s been conducting research in Madagascar in an attempt to identify the precise drivers of this seasonality. She’s found that female fruit bats in Madagascar in particular have increased viral titers during gestation and lactation, perhaps implying that female individuals are sacrificing resources that would normally be devoted to immunity to instead raise some bat babies. Some additional modeling work suggests that maternal immunity passed on to those same bat babies that wanes following the first year of life is also playing an important role in observed seroprevalence patterns. This result was dependent on fitting age-structured models to seroprevalence data from bats of different ages. How do you determine how old a bat is you might ask? Cara, the chiropteran tooth fairy, explained that age can be determined from an extracted bat tooth, and that this simple dental procedure does not affect long-term bat survival or fitness. Moving from the dental to the molecular, Cara next told us about her within-host research that seeks to explain how bats manage the dance between viral exposure, a constitutive viral immune response, and immunopathology. She’s using in vitro experiments and within-host, cell-based models to explore how viruses move between cells and between bats. Cara told us that these results suggest that bats’ constitutive immunity causes higher replication rates of viruses within individuals. It was a tour-de-force of field, modeling, and molecular work and taught everyone here at Berkeley EEID a lot about bats as reservoirs for emerging infectious diseases! Writing by Chris Hoover
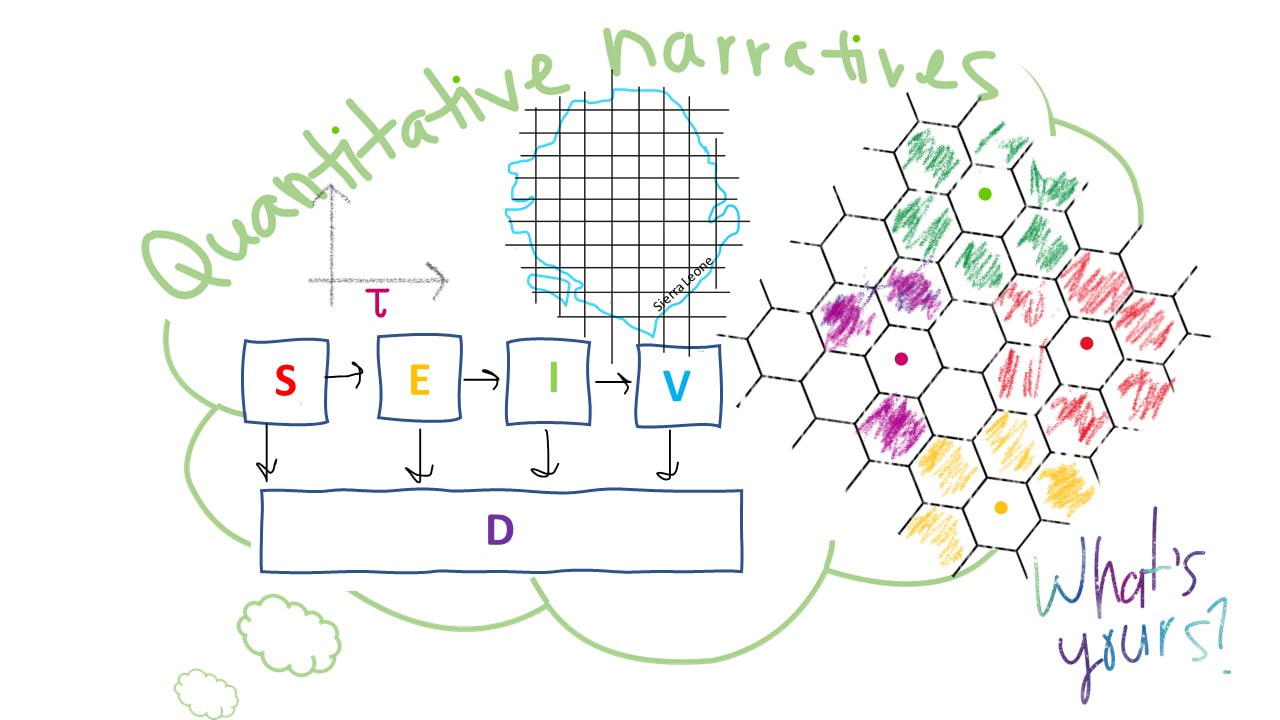 Illustration by Whitney Mgbara Science as narrative in infectious disease biology It’s been a few weeks since we’ve posted a new blog, and that delay is largely due to my tardiness—I was caught up in a grant deadline without a second to spare until now. Apologies! But a bit of time for reflection is rarely a bad thing, and it’s refreshing to revisit this talk now. On Tuesday, September 17, Professor Wayne Getz from UC Berkeley’s Department of Environmental Science, Policy, and Management presented about “Quantitative narratives in disease ecology and the recent outbreak of Ebola in Sierra Leone.” In Wayne’s own words, the talk was heavy on the “quantitative narratives” and less heavy on the Ebola, but it sure gave us plenty to think about. What is a quantitative narrative, you might ask? We don’t often think about narration as quantitative, but it certainly is. Wayne’s talk focused mostly on the fitting of mechanistic models to time series data in ecology and epidemiology, and mechanistic modeling is, by its very nature, narrative. In disease ecology, we distinguish between two broad classes of model: statistical and mechanistic. In statistical models, like the well-known linear regression, data drives the story; we plot two variables against one another and use statistical methods to demonstrate associations between those variables. In mechanistic models, we attempt to describe process, or causation, rather than correlation. This makes the approach much more active, much more narrative. All mechanistic models incorporate a process-based thesis—essentially a narrative—and we test how well each model, each narrative, recaptures the data to which we compare it. As Wayne said himself, “You can never actually prove something is true by fitting a model to data.” But if the model can convincingly recapitulate the data, we tend to accept the narrative on which the model was based. Wayne opened his talk with a broad overview of mechanistic modeling in ecology, touching on many systems that I have not reflected on much since my qualifying exam, including predator-prey models and critical community sizes for population extinction. Then, he dove into the main thread of his talk on zoonoses. Zoonotic pathogens are pathogens that are transmitted from wildlife reservoirs to human hosts, and Wayne’s research group works on several high profile zoonoses: anthrax and Ebola among them. Wayne described another two classes of model—both of the mechanistic type—which his group has used to recapitulate data from zoonotic systems. These model classes—compartmental and individual-based—represent differing approaches to narratives in disease ecology. In compartmental modeling, such as the Susceptible-Infectious-Recovered (SIR) framework, hosts are classed into broad epidemic categories, and modelers use equations to track their movements and define transitions between them. In individual-based modeling, distinct individuals are modeled explicitly, allowing for much greater flexibility in incorporating heterogeneity (i.e. in behavior or immunity) in the system. The decision over what type of model to apply to a problem is a narrative one, as different models tell different stories. My committee member in grad school, Princeton Professor Bryan Grenfell, used to say, “If you apply a complex model to a complex problem, then you have two things that you don’t understand.” Bryan was a big fan of simple compartmental models; simplifying the modeled system down to its fundamental elements tends to give the modeler much greater power to interpret the most important drivers of disease and assess how an effective intervention might be applied. Bryan also worked on a simple disease: measles, a perfectly-immunizing childhood infection that makes it easy to ignore heterogeneities between hosts because most people respond to the virus in very similar ways. In Wayne’s case, he showed us how compartmental models fit to the 2014 Ebola epidemic in West Africa failed to recapitulate the data under the simplest assumptions but did better once he incorporated a time-varying transmission rate. Only by allowing for the transmission rate to slow in the latter half of the epidemic was Wayne able to recapture these data. In his narrative, this is because human behavior changed—contact rates diminished as people stayed home out of fear. It’s a good story, but as Wayne acknowledges himself, you can never really prove it. Wayne closed by introducing us to a new platform in individual-based modeling that his lab group as developed with a software called Nova. Wayne is clearly excited about all of the gadgets embedded in the platform—it allows you to easily incorporate a within-host model (representing immune dynamics, for instance) within a compartmental model and simulate across many layers all via an accessible graphical interface. While certainly offering greater flexibility than a compartmental system, these models are a bit harder to interpret, and fitting these models to data can prove additionally challenging. Ultimately the decision over what type of model to use comes back to the research question and the story that you want to tell. If you are trying to recapitulate data with a credible narrative, then your model will need to produce its own data to compare with reality. Individual-based models hold enormous promise in application to longitudinal cohort studies where the modeler possesses data on individual hosts resampled over time—possibly even including genetic information about pathogen evolution within those hosts. In other cases, where data are less well-resolved, a compartmental approach might be more appropriate. In sum, a mechanistic model is a narrative, the disease ecologist’s way of telling a story. It’s the modeler’s challenge to find the model that is most appropriate for the narrative he or she wishes to tell. Writing by Cara Brook
Steve Parratt presents “The enemy of my enemy: the impact of a hyperparasite on a plant pathogen"9/17/2019 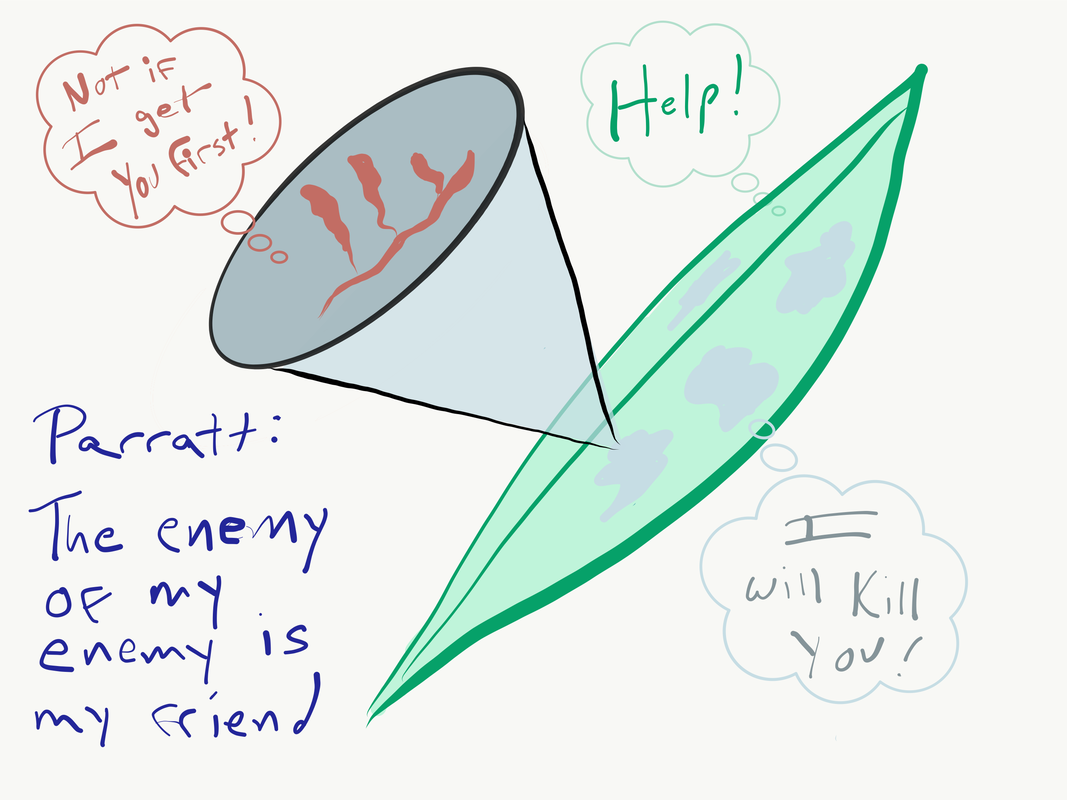 Illustration by Britt Koskella This week in the Berkeley Ecology and Evolution of Infectious Disease Seminar, we heard from Dr. Steve Parratt (University of Liverpool) on his work with hyperparasitism. Hyperparasitism, for the unfamiliar, is the phenomenon in which a parasite is itself parasitized by another organism. Hyperparasites can modify the outcome of interactions between host and parasite in unique and surprising ways. Steve started off with a quote from Jonathan Swift illustrating that people have been aware of this concept for some time.
So, naturalists observe, a flea Has smaller fleas that on him prey; And these have smaller still to bite 'em, And so proceed ad infinitum. He then introduced the two systems that he would cover, including work from his PhD and current PostDoc. The first of these projects focused on Arsenophonus nasoniae, a bacterium that parasitized the wasp Nasonia vitripennis which in turn was a parasite of Filth Flies. These bacteria are generally believed to undergo vertical transmission from mother to offspring, and have the interesting effect of killing a large proportion of the male offspring. This makes sense given their assumed mode of vertical transmission, which leaves males as a dead-end host. This also has the intentional or unintentional effect of increasing female fitness through a reduction in competition and decreased chance of inbreeding (as siblings occupy and develop on the same host). However, contrary to the assumption that vertical transmission is the primary route this parasite utilizes, Steve’s work showed that host contact and horizontal transmission were more important in the bacterium’s spread. In cases where horizontal transmission was possible the hyperparasite increased towards fixation and often caused extinction of the local population. This research calls into question the assumption that male-killing is a consequence of vertical transmission, when it may instead be due to benefits conferred during super-infection. When multiple parasite broods infect the same host (super-infection) the parasite has a higher likelihood of transmitting horizontally. In this scenario there is also more likely to be overcrowding, and so the male-killing trait may instead function simply to reduce competition between potential hosts, increasing host and personal fitness. Steve then introduced his current project; investigating the interactions between the fungal plant pathogen commonly known as Powdery Mildew and its hyper-parasite Ampelomyces quisqualis. A. quisualis is believed to negatively impact the probability of Powdery Mildew overwintering, and so Steve set out to understand the role this hyper-parasite might play in the plant-pathogen interaction. First, he showed that the hyper-parasite is variably host specific, with some strains showing evidence of local adaptation and other not. He then found that while the hyper-parasite is common at the meta-population scale it is not especially prevalent in any given population. Within a population A. quisualis seemed to be clustered within a small range of Powdery Mildew infected plants, leading to the conclusion that it is dispersal limited. When investigating the extent to which the hyper-parasite effects Powdery Mildew Steve found that it has small but significant impacts in a lab setting, but no discernible impact in natural experiments. In fact, the only time that A. quisualis had a noticeable impact of Mildew survival was during the Powdery Mildew spore stage. Finally, Steve showed that A. quisualis had a weak but significant effect during co-infection with multiple Powdery Mildew strains, but that this effect did not alter the outcome of competition. Ultimately, this research calls into question the assumption that the “enemy of my enemy is my friend”, as it seems that A. quisualis is either unable, or “un-willing” to exert a meaningful impact on the Host vs. Primary Parasite interaction. In some cases it seems that, as Steve put it: The enemy of my enemy isn’t necessarily a friend if the enemy (and consequently the hyper-parasite) do better when the host does worse. Clearly hyperparasitism is capable of impacting some host parasite interactions, but it may not be safe to assume that’s always the case. This concludes another week of the Berkeley EEID seminar, stay tuned for next week when we hear from Wayne Getz about “Quantitative narratives in disease ecology and the recent outbreak of Ebola in Sierra Leone”. Writting by Eli Mehlferber  Illustration by Nina Sokolov We kicked off the first official Berkeley Ecology and Evolution of Infectious Disease Seminar with a talk by Professor Rodrigo Almeida from ESPM on his lab’s work on Xylella fastidiosa genetics in agricultural systems. Starting with an introduction to the system, Rodrigo outlined the biology of X. fastidiosa as a multi-host, multi-vector pathogen with high rates of recombination. X. fastidiosa is transmitted by leafhopper vectors that feed on the xylem sap of many plants and can cause severe epidemics and disease in agricultural systems. It has caused large epidemics both historically and recently, so its management is hugely important to people’s livelihoods and local and regional economies. Particularly interesting to my own interests of niche breadth evolution, Rodrigo let us know that this is a highly generalist pathogen, vector, and host complex. In natural ecosystems, he proposes that X. fastidiosa populations are likely composed of many genetically diverse strains that are transmitted by multiple vector species onto multiple host plant species, leading to high levels of strain mixing and recombination. In agricultural systems, there is often one strain of X. fastidiosa being transmitted by one vector species to single host agricultural monocultures. Interestingly, there is no specificity between what bacterial strains colonize which vectors, and the vectors are generalists too, but lots of specificity between bacterial strain and host plant when it comes to disease. Getting into his lab’s recent projects, Rodrigo outlined some of the work that post-doc Andreina Castillo Siri has done to look at how the high recombination rates of X. fastidiosa is associated with disease emergence into new host plants and environments. Rodrigo showed evidence that there is lots of gene flow between introduced and local native X. fastidiosa strains at the start of novel epidemics and shared some initial hypotheses for how recombination early in epidemics may help local adaptation, leading to recombined strains spreading early in epidemics, followed by recombination slowing down as the epidemic ages and there is more ecological isolation. The lab has started to explore this idea by comparing whether X. fastidiosa strains are donors or recipients of genes from both ancestral and recent recombination events in order to identify genes that have undergone recombination and may have been important for recent epidemics. The lab has also worked on historical outbreaks of X. fastidiosa, such as one in California grapevines since the late 1800s. This epidemic spread through California quickly, and, despite few geographic barriers to gene flow, has a highly geographically structured phylogeny with local diversification. These local strains of X. fastidiosa also appear to be recalcitrant to introduction of strains from other Californian regions, suggesting that they may be adapted to their local environments. Rodrigo presented work from Mathieu Vanhove exploring the landscape genomics of X. fastidiosa and how some SNPs correlated with temperature variation. Overall, Rodrigo’s presentation was an exciting start to the seminar series and the diversity of the work he presented highlighted the overlap between the all of our research groups. After a couple of questions, we wandered over to Raleigh’s for Happy Hour to turn our focus towards beers and catching up after the summer! Writing by Elisa Visher
We've made this website to organize the various labs around campus who study the ecology and evolution of microbes and infectious disease. Research in our field is scattered across a number of departments at UC Berkeley, so by creating a weekly seminar series and happy hour, we hope to foster a collaborative community with cross-talk between all of our various expertises.
We plan to use this blog to share news, updates on our current work, summaries of what we're up to in seminar, recent papers that we've published and are reading, and blog posts about our work. Follow along to see what we're up to! |
AuthorBerkeley EEID Group Archives
February 2020
Categories
All
|
 RSS Feed
RSS Feed
